
INTRODUCTION
Higher order chromatin structure comprises of nucleosomes and DNA which further form compact structures to fit 1-2 meter DNA into a very small nucleus.
Earlier studies of chromosomes divided chromosomes divides chromosomal region into two categories : euchromatin and heterochromatin.
Heterochromatin was characterized by dense staining with a variety of dyes and a more condensed appearance, whereas euchromatin had the opposite characteristics, staining poorly with dyes and having a relatively open structure.
Heterochromatin regions of chromosomes had very limited gene expression. In contrast, euchromatic regions showed higher levels of gene expression.
Heterochromatic regions show little gene expression, but this does nnot mean that these regions are not important. In addition, heterochromatin is associated with particular chromosomal regions, including the telomere and the centromere, and is important for the function of both of these key chromosomal elements.
It is clear that DNA in both types of chromatin is packaged into nucleosomes. The difference between heterochromatin structure and euchromatin structure is how the nucleosomes in these chromosomal regions are assembled in larger assemblies.
It has become clear that heterochromatic regions are composed of nucleosomal DNA assembled into higher-order structures that result in a barrier to gene expression. In contrast euchromatic nucleosomes re found in much less organized assemblies.
HISTONE H1 BINDS TO THE LINKER DNA BETWEEN NUCLEOSOMES
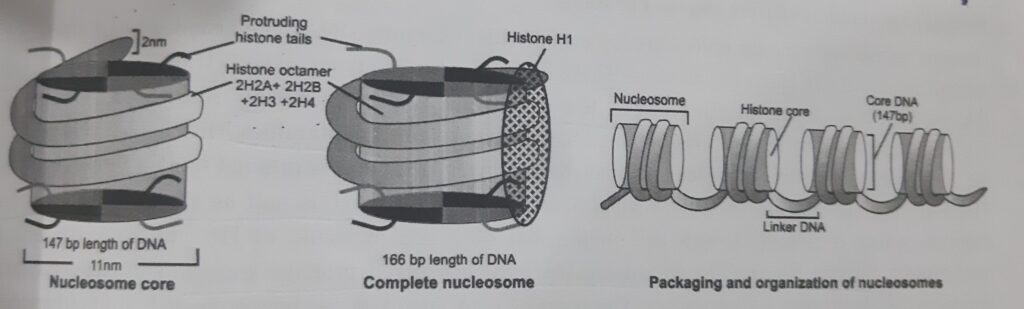
- Once nucleosomes are formed, the next step in the packaging of DNA is the binding of histone H1.
- H1, the small positively charged protein. H1 interacts with the linker DNA between nucleosomes, further tightening the association of the DNA with the nucleosome.
- Histone H1 protects DNA from action of micrococcal nuclease digestion.
- Histone H1 binds two distinct regions of the DNA duplex. Typically, these two regions are part of a single DNA molecule associated with a nucleosome.
- One of the two regions bound by H1 is the linker DNA at one end of the nucleosome. The second site of DNA binding is in the middle of the associated 147bp . Thus, the additional DNA, protected from the nuclease, is restricted to linker DNA on only one side of the nucleosome.
SECOND LEVEL OF CHROMATIN ORGANIZATION ( 30nm FIBER)

- The second level of organization is the coiling of the 11nm fiber of nucleosomes into a helical array to constitute the fiber of diameter 30nm that is found in interphase chromatin and in mitotic chromosomes. This compacts the DNA ̴̴40- fold.
- The structure of this fiber requires the histone tails and is stabilized by linker histones. There are two models for the structure of the 30nm fiber.
- In the solenoid model, the nucleosomal DNA forms a superhelix containing approximately six nucleosomes per turn.
- 11nm distance is also the approximate diameter of the nucleosome disc, suggesting that 30nm fiber is composed of nucleosome discs stacked on edge in the form of a helix.
- The linker DNA circles around the central axis as the DNA moves from one nucleosome to the next.
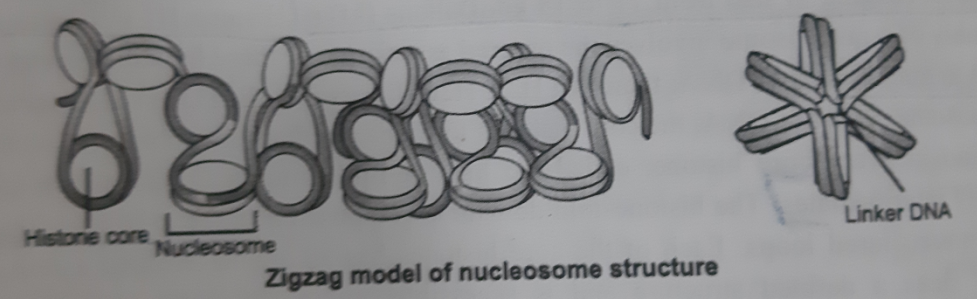
- Alternative of this model is “Zigzag” model . In this model zigzag pattern is formed upon H1 addition. The 30nm fiber is a compacted form of these zigzag nucleosome arrays.
- Zigzag model requires the linker DNA to pass through the central axis of the fiber in a relatively straight form.
- Both forms of the 30nm fiber may be found in cells depending on the local linker DNA length.
NUCLEAR SCAFFOLD (COMPOSED OF NON-HISTONE CHROMOSOMAL PROTEINS)
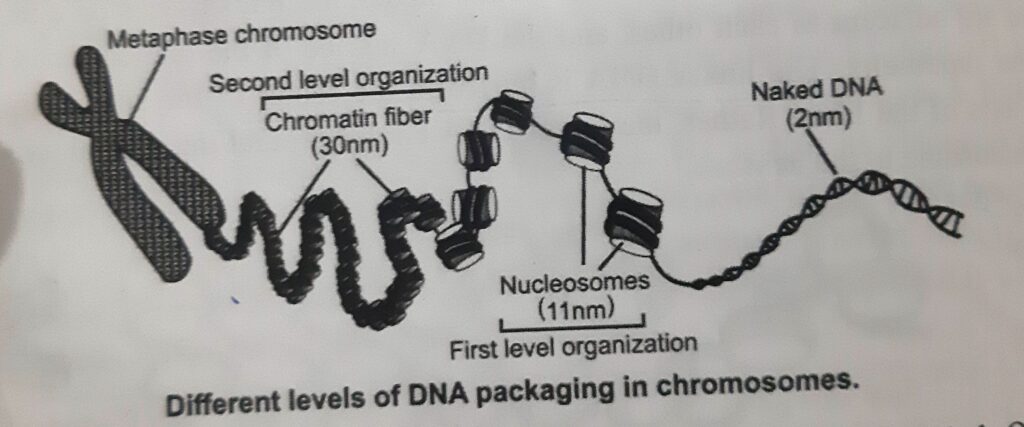
- The packaging of DNA into nucleosomes and the 30nm fiber results in the compaction of the linear length of DNA by ̴40-fold. This is insufficient to fit 1-2 m of DNA into nucleus ̴10-5 m across.
- Compaction of 30nm fiber is required to compact the DNA further.
- The 30nm fiber is then further folded and compacted into interphase chromatin or into mitotic chromosomes. This results in ̴1000-fold compaction in euchromatin, interchangeable with packing into mitotic chromosomes to achieve an overall ̴ 10,000 fold compaction.
- Two classes of proteins that contribute to the nuclear scaffold have been identified.
- One of these is topoisomerase II (Topo II), which is abundant in both scaffold preparations and purifieed mitotic chromosomes. Treating cells with drugs that results in DNA breaks at the sites of Topo II DNA binding generates DNA fragments that are 50 kb in size.
The second class of proteins are SMC Proteins ( cohesin and condensin) which are also abundant components of the nuclear scaffold. The association of these proteins with the nuclear scaffold may serve to enhance their functions by providing an underlying foundation for their interactions with chromosomal DNA.
Share this:

Discover more from ZOOLOGYTALKS
Subscribe to get the latest posts sent to your email.